Abstract: Lipid peroxidation processes occurring in tissues and involved in physiological and biochemical processes are very important in the life activity of hydrobionts. As a result of interaction of free radicals with organic molecules, new radicals are formed, which continue chain reactions. Peroxidation is a chain and multistage process. Such factors as radiation, UV and electromagnetic radiation, heavy metals, xenobiotics, and pharmaceuticals significantly influence the acceleration of free radical and peroxide processes in tissues of living organisms.
Keywords: lipid metabolism parameters, lipid peroxidation, freshwater hydrobionts.
Введение. Процесс перекисного окисления липидов протекает в три стадии: инициация, развитие и обрыв цепи. Инициация последовательного окисления полиненасыщенных жирных кислот клеточной мембраны в перекисное соединение происходит в результате свободнорадикальной атаки активных форм кислорода, обладающей достаточной реактивностью для отщепления атома водорода от метиленовой (-СН2 — ) группы. Так как атом водорода имеет только один электрон, при отделении Н˙ неспаренный электрон остается на углероде (-СН-). В биологических мембранах происходит отрыв атома водорода в α-положении по отношению к двойной связи у полиненасыщенных жирных кислот, входящих в состав фосфолипидов. Наличие двойной связи в жирной кислоте ослабляет связь С-Н в смежных углеродных атомах и тем самым облегчает отщепление Н˙. Чем длиннее ненасыщенная боковая цепь жирной кислоты, тем сильнее она может подвергаться перекисному окислению [1]. Радикал с углеродом в центре претерпевает молекулярную перегруппировку с образованием диена, содержащего сопряженные двойные связи, который в дальнейшем соединяется с кислородом и образует радикал пероксила, способный отделить атом водорода от другой жирной кислоты, начиная, таким образом, цепную реакцию. Возникающие свободные радикалы кислорода атакуют остатки полиненасыщенных жирных кислот в липидах с образованием жирнокислотных радикалов, которые, взаимодействуя с молекулярным кислородом, превращаются в перекисные радикалы. Этот перекисный радикал органического соединения в дальнейшем взаимодействует с новыми остатками жирных кислот с образованием гидроперекиси и новых радикалов[2, 3].
Перекисное окисление липидов представляет собой процесс, связанный с активацией кислорода, особенность которого заключается в том, что молекула кислорода присоединяется к свободному радикалу. В результате образуется новый радикал – перекисный радикал органического соединения (гидропероксид). Далее происходит взаимодействие этого радикала с новой молекулой органического соединения.
К продуктам цепной реакции перекисного окисления липидов относятся разновидности гидроперекисей и циклических перекисей. Гидроперекиси относят к первичным продуктам перекисного окисления липидов. При распаде гидроперекисей наряду с ионом гидроксила образуется алкоксильный радикал, разрушающий структуру молекулы жирной кислоты на фрагменты разной величины. При трансформации пероксидов полиненасыщенных жирных кислот образуются вторичные продукты перекисного окисления липидов, к которым относится малоновый диальдегид [6].
Цель работы – исследовать динамику ключевых показателей липидного обмена и перекисного окисления липидов в тканях легочных пресноводных моллюсков в зависимости от сезона года и местообитания.
Методы и материалы. При проведении исследований использовались легочные пресноводные моллюски – прудовик обыкновенный (Lymnaea stagnalis) 108 особей. Моллюски собирались из водоемов четырех районов Витебской области (таблица 1). В каждой исследовательской подгруппе содержалось по 9 моллюсков. Сбор осуществлялся в осенний (октябрь), весенний (апрель) и летний (июль) сезоны. Особи собирались вручную.
Таблица 1
Места сбора моллюсков
| Район сбора моллюсков | Место сбора | Название водоема |
| Витебский р-н | г. Витебск | р. Витьба |
| Дубровенский р-н | д. Шеки | оз. Афанасьевское |
| Ушачский р-н | д. Дубровка | оз. Дубровское |
| Шумилинский р-н | а/г Башни | оз. Будовесть |
Определение содержания общего холестерола и триглицеридов в гемолимфе легочных моллюсков проводили спектрофотометрически [5]. У моллюсков количественное установление продуктов перекисного окисления липидов (ТБК-ПВ) проводили по методу Uchiyama и Mihara [6], определение ГSН проводили по методу Beutler [7]. Активность каталазы определяли по реакции с молибдатом аммония [8]. Математическую обработку полученных результатов проводили методами параметрической и непараметрической статистики с использованием пакета статистических программ Microsoft Excel 2012, STATISTICA 6.0.
Эксперимент. Для объетивного биомониторинга с использованием живых объектов необходимо учитывать сезонные изменения в ключевых показателях метаболизма, поэтому актуальным является определение динамики показателей липидного обмена и перекислого окисления липидов у моллюсков с последующей оценкой различий в антропогенном влиянии, обусловленных местом обитания.
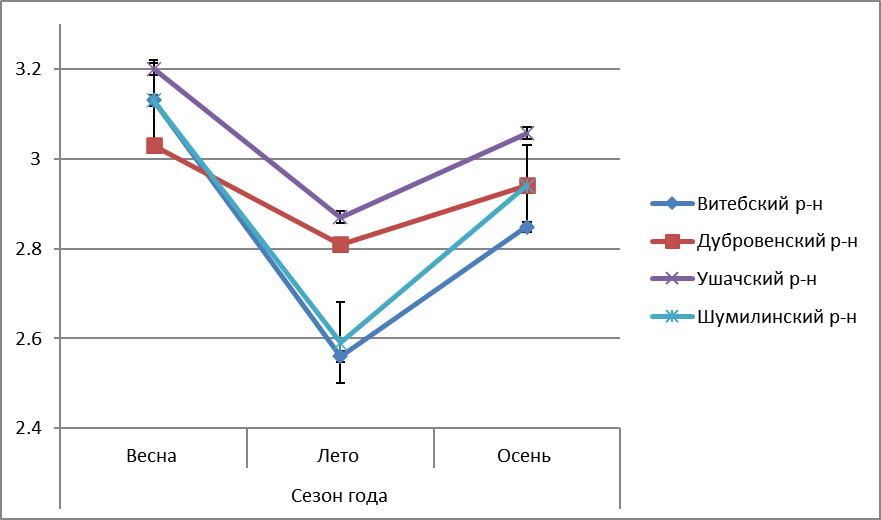
Моллюски, обитающие в водоёмах Витебской области, характеризовались следующей динамикой содержания общего холестерола, триглицеридов в гепатопанкреасе, продуктов перекисного окисления липидов в зависимости от времени года, местообитания (рисунки 1-5).
|
|
|
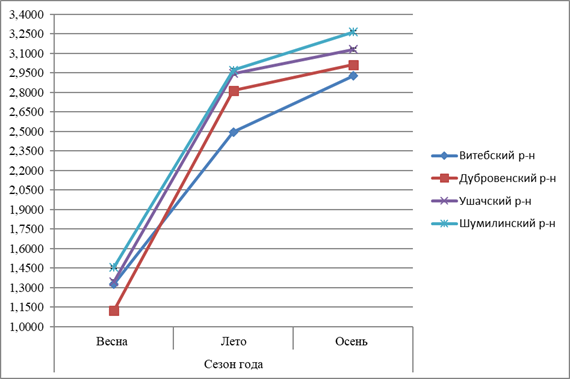
Рисунок 1 – Сезонная динамика содержания общего холестерола в гепатопанкреасе L. stagnalis, обитающих в водоёмах Витебской области |
|
|
|
Рисунок 2 – Сезонная динамика содержания триглицеридов в гепатопанкреасе L. stagnalis, обитающих в водоёмах Витебской области |
 |
|
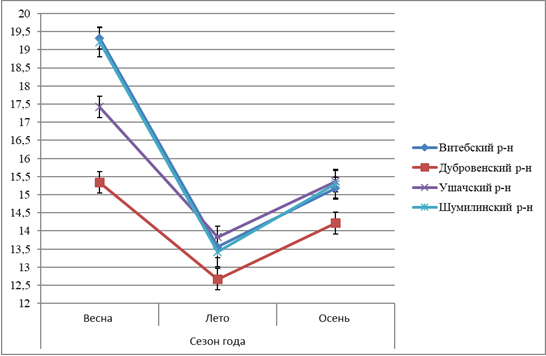
Рисунок 3 – Сезонная динамика содержания ТБК-ПВ в гепатопанкреасе L. stagnalis, обитающих в водоёмах Витебской области |
|
|
|
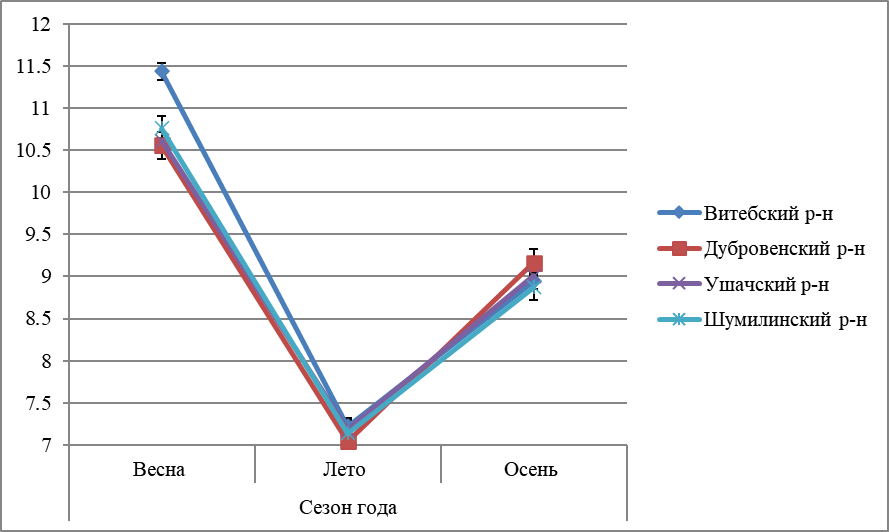
Рисунок 4 – Сезонная динамика активности каталазы в гепатопанкреасе L. stagnalis, обитающих в водоёмах Витебской области |
 |
|
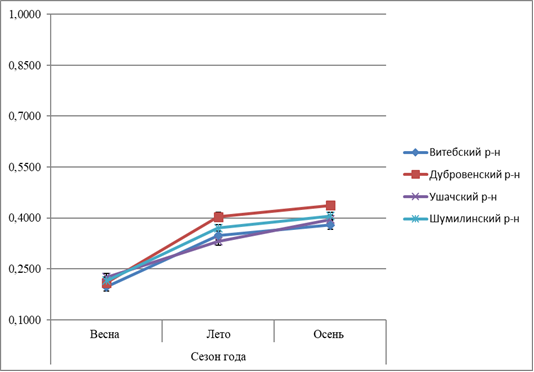
Рисунок 5 – Сезонная динамика содержания восстановленного глутатиона в гепатопанкреасе L. stagnalis, обитающих в водоёмах Витебской области |
Особенности липидного обмена легочных пресноводных моллюсков связаны c физиологическими особенностями, а также с экологическими характеристиками исследуемых водоёмов и их прибрежных зон.
Содержание ОХС и ТГ у прудовика обыкновенного имеет сходную сезонную динамику: наименьший уровень в весенний период, что связано с неблагоприятными природно-экологическими условиями для жизнедеятельности, более высокие летом и осенью (рисунки 1-2).
Река Витьба имеет выраженные признаки антропогенного воздействия. Наличие на ее берегах зон отдыха усугубляет это воздействие. На берегах и в воде реки можно наблюдать много мусора. На дне реки обнаружен черный ил, что свидетельствует о большом количестве органических веществ в воде. Вода имеет желтоватый оттенок и легкий болотный запах, что является начальными признаками эвтрофикации водоема.
Озеро Афанасьевское Дубровенского района подвергается сильной антропогненной нагрузке, так как используется для мелиорации земель, что приводит к загрязнению воды и береговой зоны водоема.
Озеро Дубровское находятся относительно далеко от крупных промышленных центров и крупных автомагистралей, отличается чистой водой, лишенной вредных примесей.
Озеро Будовесть Шумилинского района подвергается слабой антропогенной нагрузке, т.к. не используется в промышленных и сельскохозяйственных целях и в него не осуществляется сброс сточных вод.
Результаты, представленные на рисунках 3-5, показывают, что сезонные изменения условий окружающей среды оказывают влияние на антиоксидатную систему легочных моллюсков, приводят к активации процессов перекисного окисления липидов в весенний и осенний периоды года, что доказывается увеличением содержания ТБК-ПВ во всех экспериментальных группах. Изменения содержания ТБК-ПВ имеют сезонный характер. Установлено, что наибольшее содержание данного показателя фиксируется в весенний период, наименьшие значения – в летний период сбора моллюсков. Полученные изменения в концентрации ТБК-ПВ имеют однотипный характер во всех исследуемых районах сбора моллюсков: самое высокое значение в весенний период, среднее значение в осенний период, наименьшее значение в летний период (рисунок 3).
Активность каталазы гепатопанкреаса Lymnaea stagnalis также имела сезонный характер изменений (рисунок 4). Выявлено: активность каталазы выше весной, что связано с усилением неблагоприятного воздействия факторов внешней среды обитания и возрастанием окислительного стресса. Динамика изменений активности каталазы уменьшается в последовательности весна → осень → лето, что свидетельствует о повышении активности антиоксидантной системы в весеннее время года из-за необходимости в утилизации большего количества пероксид водорода, образующегося при активации окислительных процессов.
В летной период сбора у моллюсков содержание восстановленного глутатиона меньше, чем весной и осенью (рисунок 5). Установлено: содержание восстановленного глутатиона в летнее время имеет самые низкие показатели, т.к. в это время степень неблагоприятного воздействия факторов окружающей среды минимальна. Показатели в весеннее и осеннее время превышают значения в летнее время сбора. Однако весной вследствие низкой температуры и недостатка пищи моллюски испытывают стресс и значение показателей выше, чем в осеннее время сбора.
Результаты. Результаты проведенных исследований позволили сделать следующие выводы:
1) изменения показателей липидного обмена L. stagnalis, обитающих в природных водоёмах Витебской области характеризуются следующими сезонной динамикой: содержание общего холестерола и триглицеридов у прудовика обыкновенного в весенний период ниже по сравнению с содержанием данных показателей в гепатопанкреасе в летний и осенний периоды;
2) установлено, что уровни ТБК-ПВ в гепатопанкреасе моллюсков изменяются однотипно во всех исследуемых водоемах: самые низкие значения летом. Содержание ТБК-ПВ оказалось более высоким в проточных водоемах. При исследовании активности каталазы в гепатопанкреасе моллюсков выявлена аналогичная сезонная динамика: Содержание восстановленного глутатиона в гепатопанкреасе моллюсков изменяется аналогично.
3) при проведении системно-экологического анализа изменений биохимических показателей Lymnaea stagnalis было выявлено, что сезон года и место обитания оказывают существенное воздействие на обмен веществ у пресноводных моллюсков, что ускоряют или замедляют процессы жизнедеятельности данных организмов. Были изучены биохимические показатели моллюсков из следующих водоемов Витебской области: р. Витьба (г.Витебск), оз. Афанасьевское (д. Шеки), оз. Будовесть (а/г Башни), оз. Дубровское (д. Дубровка). Наиболее чистыми водоемами, обладающими более благоприятной средой для обитания изучаемых организмов, являются озера Будовесть, Дубровское и Афанасьевское, а менее пригодным для жизни является река Витьба.
References
1. Зенков, Н. К. Окислительный стресс / Н.К. Зенков, В. З. Ланкин, Е. Б. Меньшикова. – Москва: Наука, 2001. – 342 с.2. Осипов, А.Н. Активные формы кислорода и их роль в организме / А.Н. Осипов, О.А. Азизова, Ю.А. Владимиров // Успехи биол. химии. – 1990. – Т. 31. – С. 180-208.
3. Bielski, B.H. A study of the reactivity of HO2/O2 – with unsaturated fatty acids / B.H. Bielski, R.L. Arudi, M.W. Sutherland // J. Biol. Chem. – 1983. – Vol. 258. – P. 4759-4761.
4. Ланкин, В. З. Метаболизм липоперекисей в тканях млекопитающих / В. З. Ланкин // Биохимия липидов и их роль в обмене веществ : [сб. ст.] / АН СССР, Науч. совет по пробл. биохимии им. М. М. Шемякина ; отв. ред. С. Е. Северин. – Москва: Наука, 1981. – С. 75-95.
5. Чиркин, А.А. Липидный обмен / А.А. Чиркин [и др.] // Медицинская литература. – М., 2003. – 122с.
6. Uchiyama, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. / М. Uchiyama, M. Mihara. // Analit. Biochem.. – 1978.– V. 86. – P. 271–278.
7. Beutler, E. Red cell metabolism a manual of biochemical methods / E. Beutler. – Orlando: Grune & Stration, 1990. – P. 131–134.
8. Королюк, М.А. Метод определения активности каталазы / М.А. Королюк [и др.] // Лаб. Дело. - 1988. – № 1. – С. 16–19.