Abstract: Actuality of study of descriptions of arboreal plants for planting and protective setting allows objectively to estimate their priorities in relation to mechanical composition, humidity and vozduhorazdelitelnoj wood. The selection of arboreal plants on the basis of analysis of their hydrophysical and hydrochemical properties will allow to promote biological stability of planting, increase a their life span, execute protective functions, change the morphological look of trees and stands.
Keywords: limiting factors, thermodynamics gradient, hydrophysical properties, isotherm of capillary evaporation
Эффективность подбора древесных растений в условиях засушливой степи и полупустыни зависит от таких особенностей пород древесины, как засухоустойчивость, морозостойкость и сравнительная долговечность. Отсутствие данных особенностей у древесной породы ограничивает возможность ее применения, Продуктивность и биологическое разнообразие связаны с несоответствием ассортимента древесных растений условиям роста.
При создании биологически устойчивых защитных лесных насаждений в лесорастительных условиях изучается функциональная влажность древесных растений, так как водный стресс замедляет физиологические процессы. Оптимальные условия фотосинтеза создаются только при постоянной влажности ассимиляционного аппарата. Для успешного процесса фотосинтеза необходимо стабильное увлажнение [1]. К лимитирующим факторам выращивания ассортимента древесных растений относят:
— поглощение воды и восходящий ток;
— водный дефицит и физиологические процессы;
— водный режим растительной клетки. Важными двигателями влагопереноса являются:
— корневое давление, как нижний концевой двигатель, который не превышает нормы от 1 до 10 атм;
— сосущая сила атмосферы, одинаковая для всех пород, произрастающих на одной территории.
Так как термодинамический градиент на линии «почва — древесное растение» по величине меньше, чем на линии «древесное растение — атмосфера», то для устойчивого роста древесных пород в степи насаждения должны получать дополнительное увлажнение. Или же их древесина должна обладать низким потенциалом влагопереноса, чтобы длительное время противостоять огромной сосущей силе атмосферы. Установлено, что у древесины удельная поверхность больше, чем у почвы, что уменьшает существующий градиент на линии «древесное растение — атмосфера».
Таким образом, потенциал влагопереноса служит критерием засухоустойчивости и двигателем влагопереноса. Этот фактор объясняет недостаток или избыток влаги в почве, атмосфере и древесине. Процессы фотосинтеза и дыхания в древесине и почве, как в сложной окислительно-восстановительной системе, связаны с реакцией окисления и восстановления, и ее потенциал зависит от рН раствора [2].
Возможности влагопереноса свежесрубленной древесины зависит от вида и влажности древесного растения. Неравномерность увлажнения отрицательно сказывается на жизни растений степной и полупустынной зонах. Для этих зон характерна неравномерная сумма осадков за год, низкая влажность воздуха и высокая испаряемость. Неблагоприятный температурный режим не влияет на ассортимент деревьев и кустарников для защитного лесоразведения в аридной зоне, включает сто двадцать пород и пополняется другими видами.
По изученным данным дефицит влаги в почве и атмосфере рассматривается в отрыве от влагообеспеченности древесных растений. Гидрофизические константы древесины не зависят от вида древесной породы, так как химический состав древесного вещества для всех пород приблизительно одинаков.
Опытным путем установлено, что гидрофизические свойства живой древесины видоспецифичны. В изотермических условиях, если влажность воздуха 76% и более, то равновесная влажность древесины разных пород различная.
Если влажность воздуха меньше 76 % , то показатели между породами схожи с гидрофизическими свойствами сухой древесины. Если за влагоемкость древесины и листьев принимать их влажность после полного насыщения образцов под вакуумом до конца барботирования, то лимиты влажности древесины можно рассчитать по аналогии с данными для почвы:
— благоприятная — 0,8…0,9;
— достаточная — 0,6…0,8;
— напряженная — 0,5 … 0,6;
— критическая — 0,3 … 0,5.
Пробные площади деревьев и модели на их рост производятся общепризнанными способами. Отбор образцов древесины для изучения гидрофизических свойств можно производить из свежесрубленных здоровых деревьев I–II классов Крафта [3].
В системе «дерево — среда» имеются физические характеристики почвы, а гидрофизические свойства живой древесины основных лесообразующих пород видоспецифичны. Изотерма капиллярного испарения дуба и березы, заболони и ядра существенно различаются (таблица 1).
Таблица 1.
Изотерма капиллярного испарения древесины
(в числителе – заболонь, в знаменателе — ядро)
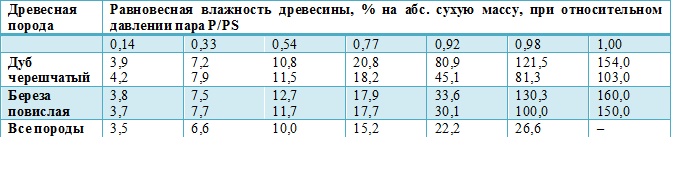
Примечание. Р, РS – давление пара соответственно при данной влажности воздуха и полном насыщении воздуха влагой.
Изотерма капиллярного испарения, как фундаментальная гидрофизическая характеристика древесины, является потенциалом влагопереноса в диапазоне ее влагосодержания, что определяет древесину как видоспецифический двигатель влагопереноса: сосущая сила атмосферы одинакова для всех, а корневое давление на порядок меньше (таблица 2).
Таблица 2.
Взаимосвязь физических показателей древесины главных пород
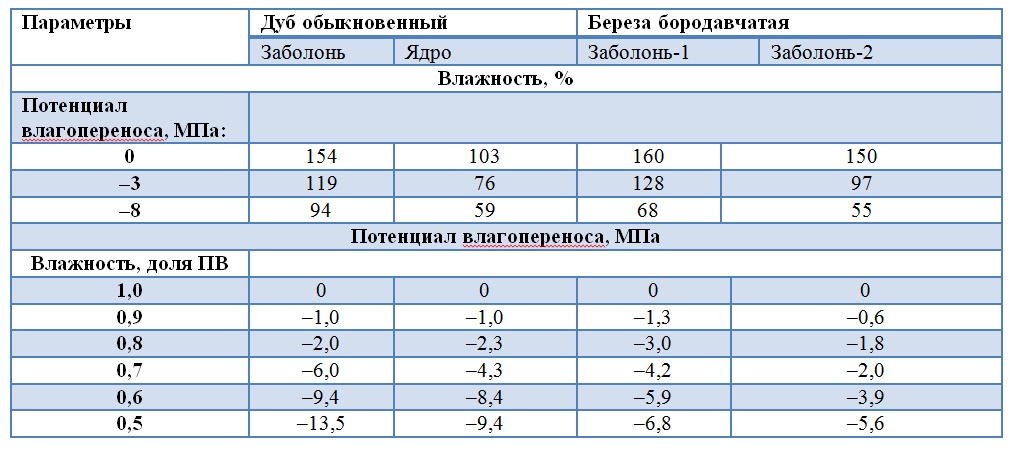
Древесина ствола, как резервуар, демпфер и проводник влаги, хранит запасы питательных веществ, в засушливую погоду из них образуется катаболическая вода [4].
Древесные породы по засухоустойчивости можно классифицировать по значению потенциала влагопереноса древесины при влажности 0,5 (рисунок 1).
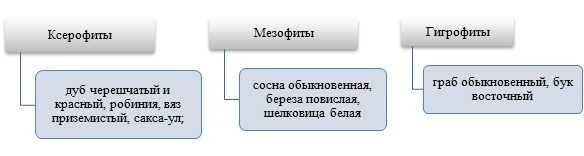
Рисунок 1. Классификация древесных пород по засухоустойчивости
По градиенту давления влаги в древесине определяется направление влагопереноса. Ядро некоторых пород деревьев в диапазонах влажности становится акцептором влаги. Это ограничивает их применение в жестких растительных условиях леса. Но породы-доноры по функции ядра влагообменны. Заболонь дуба в течение всей вегетации имеет более низкий потенциал влагопереноса по сравнению с ядром, даже когда различия по влажности древесины между ними составляют 25 … 30 %. Гидрофизические характеристики дуба объясняются отнесенностью дубрав к почвам тяжелого механического состава.
По изотерме капиллярного испарения рассчитываются лимиты функциональной влажности древесины и листьев, которые служат основой экспресс-анализа и прогноза состояния древесных пород.
Одним из основных параметров подбора древесных растений в условиях засушливой степи и полупустыни является фундаментальная математическая доказанность зависимости фотосинтеза от оводненности ассимиляционных тканей [1]. Колебания влажности листьев, побегов, древесины в течение суток, месяцев, сезонов года в степи, лесостепи и бореальной зоне существенны. Увеличение площади питания снизит колебания влажности, эффективность фотосинтеза до определенных лимитов. Площадь питания определяют по диаметру крон средних деревьев, равному в устойчивых насаждениях 50 % длины ствола [5].
Устойчивые древостои состоят преимущественно из деревьев I и II классов Крафта, так как при интенсивных рубках в один-два приема выбирают деревья низших классов и оставляют только деревья будущего. Любой довод о преимуществах господствующих деревьев по сравнению с деревьями III–V классов Крафта свидетельствует в пользу устойчивых древостоев. К этим особенности относят:
— влажность древесины;
— масса, влажность и площадь хвои;
— число шишек;
— объем среднего дерева;
— размер ядра;
— размер и объем крон;
— величину КОП;
— динамике рангов;
— смолопродуктивность;
— число здоровых деревьев;
— снижение прироста в засуху;
— отзывчивость на рубки.
Могучие вековые деревья на опушках при распаде многих видов защитных насаждений степи свидетельствует об упущенных возможностях по созданию устойчивых лесов (80 % здоровых деревьев растет на опушках) [6].
Таким образом, приемы формирования заведомо устойчивых деревьев состоят в эффективной профилактике этиолированности древостоев, стабилизации водного режима ассимиляционного аппарата для обеспечения фотосинтеза. Для создания многоцелевых устойчивых лесов необходимо увеличивать запасы доступной влаги в почве и древесине деревьев будущего, существенно увеличивать размеры крона и количество запасов питания для получения катаболической влаги. Древесных видов пород с морфологически одинаковой древесиной и ее анатомической структурой, влажностью, плотностью, цветом, запахом не существует. Потенциал влагопереноса древесины в системе «почва — древесное растение – атмосфера» является двигателем переноса влаги. Он служит критерием засухоустойчивости и рассчитывается по изотерме капиллярного испарения.
References
1. Берри Д.А. Зависимость фотосинтеза от факторов окружающей среды / Фотосинтез. – М.: Мир - Т. 2. - С. 273–364.2. Казарян В.О. Физиологические аспекты эволюции от древесных к травам. Л.: Наука.- 348 с.
3. Крафтс А. Вода и ее значение в жизни растений / А. Крафтс, Х. Карриер, К. Стокинг. – М.: ИЛ. - 388 с.
4. Густова, А. И. Изучение свойств свежесрубленной древесины для оценки устойчивости в условиях степной зоны / А. И. Густова // Лес, Наука, Молодежь: сб. науч. тр. / Воронежская государственная лесотехническая академия. Воронеж, 2006. - С. 37-42.